ЭКСТРАПИРАМИДНАЯ СИСТЕМА (systema extrapyramidale) - система ядер головного мозга и двигательных внепирамидных (экстрапирамидных) проводящих путей, осуществляющая непроизвольную, автоматическую регуляцию и координацию сложных двигательных актов, регуляцию мышечного тонуса, поддержание позы, организацию двигательных проявлений эмоций.
Экстрапирамидная система в отличие от пирамидной системы (см.) не является строго очерченной анатомической и функциональной системой. Она объединяет некоторые отделы коры головного мозга (см.), базальные ядра (см.), ядерные образования мозгового ствола (см. Головной мозг), мозжечок (см.), сегментарный аппарат спинного мозга (см.), а также обширные коммуникации, осуществляющие мгновенную функциональную интеграцию многих нейрональных систем, обеспечивающих сложную организацию двигательных и поведенческих актов.
Анатомия
Экстрапирамидная система включает полосатое тело (corpus striatum), состоящее из хвостатого ядра (nucleus caudatus) и чечевицеобразного ядра (nucleus lenticularis), медиальные ядра таламуса (nuclei mediales thalami), субталамическое ядро (nucleus subthalamicus, s. corpus Luysi), ядра гипоталамуса (nuclei hypothalamici), черное вещество (substantia nigra), красное ядро (nucleus ruber), ядра ретикулярной формации (nuclei formationis reticularis), оливу (oliva) продолговатого мозга. Хвостатое ядро состоит из головки (caput), тела (corpus) и хвоста (cauda); чечевицеобразное ядро - из скорлупы (putamen), бледного шара (globus pallidus), медиальной и латеральной мозговых пластинок (laminae medullares medialis et lateralis). В экстрапирамидную систему входят также двигательные экстрапирамидные проводящие пути: корковые пути, берущие начало от нейронов коркового двигательного поля 4 и нейронов, расположенных в соматосенсорных корковых полях, связывающие кору полушарий головного мозга с образованиями экстрапирамидной системы; стриопаллидарные пути, соединяющие образования экстрапирамидной системы между собой; трункоспипальные пути, идущие от перечисленных двигательных ядер головного мозга к двигательным ядрам спинного мозга и черепно-мозговых нервов. К экстрапирамидной системе относят и мозжечок.
Экстрапирамидная система представляет собой филогенетически старую систему. Конечный мозг низших позвоночных не имеет коры, а клеточные скопления, образующие базальные ядра, залегают в его глубине. Экстрапирамидная система низших позвоночных является высшим отделом, принимающим сигналы от органных рецепторов и посылающим импульсы к мышцам через центры спинного мозга. У рыб из образований экстрапирамидной системы имеется только бледный шар чечевицеобразного ядра, у амфибий появляется скорлупа этого ядра. У рептилий и птиц с развитой корой головного мозга образуются новые базальные ядра (например, хвостатое ядро), но сохраняются непрямые нисходящие пути от этих ядер. В то же время прямая связь между корой головного мозга и спинным мозгом у них отсутствует. Только у млекопитающих помимо экстра пирамидных появляются и прямые нисходящие пирамидные двигательные пути от коры головного мозга к двигательным центрам спинного мозга.
На основании данных о развитии базальных ядер в экстрапирамидной системе выделяют ее ядерную часть - стриопаллидарную систему, при этом хвостатое ядро и скорлупу чечевицеобразного ядра объединяют под названием «стриатум» (striatum), или «неостриатум», а бледный шар обозначают как паллидум (pallidum). К системе паллидума относят также черное вещество и красное ядро (см. Средний мозг). Стриатум - филогенетически более молодое образование, чем паллидум. Образования стриопаллидарной системы связаны между собой стриопаллидарными проводящими путями (см. Двигательные центры, пути).
Физиология
Основные физиологические функции экстрапирамидной системы обеспечивают координацию двигательных актов человека и животных, регуляцию мышечного тонуса и поддержание позы, организацию двигательных проявлений эмоций. Сложность строения экстрапирамидной системы, обширность связей ее структур с различными образованиями головного мозга делают трудным понимание физиологических механизмов экстрапирамидной регуляции двигательных актов. В отличие от пирамидной системы (см.) экстрапирамидная система не разделяется па отдельные пути, а представляет собой сложную систему двигательных ядер и связей между ними, а также связей двигательных центров различных функциональных уровней головного мозга с эфферентными нейронами спинного мозга (см.) и ядрами черепно-мозговых нервов через многочисленные подкорковые и стволовые структуры. В спинном мозге импульсы, поступающие по нисходящим пирамидному тракту и волокнам экстрапирамидной системы, взаимодействуют с возбуждениями, приходящими по афферентным путям от пропроирецепторов. Процесс интеграции возбуждений на уровне спинного мозга является важным звеном в механизме не только произвольных, но и непроизвольных. Начало экстрапирамидной системы дают преимущественно аксоны нейронов коркового двигательного поля 4, а также нейроны, расположенные в соматосенсорной коре. Вместе с тем многие волокна экстрапирамидной системы начинаются и в других сенсорных областях коры (слуховой, зрительной и вкусовой) и в ассоциативных зонах лобной, теменной и височных долей мозга (см. Кора головного мозга). От клеток коры головного мозга импульсы по экстралирамидным путям направляются к нейронам таламуса (см.), гипоталамуса (см.), моста головного мозга (см.), красных ядер, черного вещества и ретикулярной формации (см.). От поля 4 экстрапирамидные волокна вместе с волокнами пирамидного тракта идут в составе внутренней капсулы к наиболее древним группам двигательных ядер - базальным ядрам (см.). Эти ядра занимают центральное место среди структур экстрапирамидной системы, являясь высшим надсегментарным аппаратом, обеспечивающим регуляцию двигательных актов с участием различных мышечных групп. С помощью базальных ядер осуществляются все синергии, входящие в состав таких сложных двигательных актов, как ходьба, бег, лазанье и др. При участии экстрапирамидной системы создается плавность движений и устанавливается исходная поза для их выполнения. Экспериментальные исследования и клинические наблюдения позволяют отметить многообразие форм и в то же время однозначность (усиление или подавление) двигательных реакций при разрушении или раздражении экстрапирамидных образований. Повреждения образований экстрапирамидной системы вызывают различные нарушения в двигательной сфере. Так, при повреждении внутренней капсулы в глубине полушария головного мозга, где проходят как пирамидные, так и экстрапирамидные волокна, наблюдается спастическое повышение тонуса мышц парализованных конечностей (см. Гемиплегия), которое обусловлено разрушением именно экстрапирамидных проводников, оказывающих в норме тормозное влияние на ретикулярную формацию. Обычно при изолированном повреждении пирамидного тракта на уровне продолговатого мозга (см.), где большинство экстрапирамидных волокон идет отдельно и не затрагивается при повреждении, наблюдается мышечная гипотония. Наиболее выраженные тормозные эффeкты проявляются в эксперименте при стимуляции экстрапирамидных областей коры головного мозга. Как показала Тауэр (S. S. Tower, 1940), корковое торможение может быть двух видов - торможение мышечного тонуса (см.) и торможение совершающегося движения (см.). Последний вид торможения характерен для нисходящих корковых экстрапирамидных влияний, они играют важную роль в регуляции физиологической активности двигательных экстрапирамидных ядер мозгового ствола, от которых к двигательным центрам спинного мозга поступает мощный поток возбуждений. Прямые пирамидные корковые связи ускоряют движения и обеспечивают возможность их более тонкой дифференцировки. Экстрапирамидные движения, вызываемые стимуляцией различных участков коры, более медленны и стереотипны. В отсутствие тормозных влияний коры головного мозга движения превращаются в ряд судорожных мышечных сокращений.
Среди структурных образований экстрапирамидной системы стриатум считается высшим подкорковым регуляторно-координационным центром организации движений, в то время как паллидум, влияя на нейроны спинного мозга через структуры среднего и продолговатого мозга, координирует тонус и фазовую двигательную активность мышц. Деятельность стрио-паллидарных структур связана с выполнением медленных сложных движений, таких как медленная ходьба, перешагивание через препятствие, вдевание нитки в иголку и др. При осуществлении какого-либо движения в ограниченном или в чрезмерном объеме афферентная обратная снизь от проприоцепторов сигнализирует об этом, и от базальных ядер к двигательным областям коры головного мозга и к стволовым структурам поступают сигналы коррекции. Таким образом, пирамидная и экстрапирамидная системы успевают по ходу выполнения движений внести исправления в непрерывный поток двигательных возбуждений. После разрушения стриатума возникают непроизвольные хаотические движения отдельных конечностей - гиперкинезы (см.). Нарушения экстрапирамидной регуляции произвольной и непроизвольной двигательной активности мимических мышц (см. Мимика) приводят к неадекватному внешнему выражению эмоций (см.), непроизвольному смеху и плачу или полному отсутствию мимического выражения (маскообразное лицо).
Одной из функций бледного шара является торможение нижележащих ядер среднего мозга. При повреждении бледного шара наблюдается увеличение тонуса скелетной мускулатуры (гипертонус) вследствие освобождения красного ядра среднего мозга от тормозящего влияния паллидума. Раздражение бледного шара приводит к повышению тонуса мышц и тремору конечностей, а также к ограничению и скованности движений. Эти явления устраняются при разрушении паллидума. Торможение движений наблюдается и при раздражении полосатого тела. Подобные эффекты подавления движений отмечаются при раздражении так наз. тормозных зон экстра пирамидной системы (поясная кора головного мозга, части моторной коры, хвостатое ядро, мозжечок, ретикулярная формация).
Стриатум дает начало многим двигательным путям экстрапирамидной системы, среди которых выделяют эффекторный путь, идущий к паллидуму, а далее через красное ядро и руброспинальный тракт - к спинному мозгу. Стриатум, включающий хвостатое ядро и скорлупу чечевицеобразного ядра, достаточно хорошо морфологически и нейрофизиологически изучен. Обилие внутренних связей является особенностью его строения, хотя небольшое число нейронов посылает свои аксоны и к другим структурам мозга, в том числе к ядрам таламуса и гипоталамуса. Экспериментальные физиологические исследования с применением микроэлектродов показали (см. Микроэлектродный метод исследования), что стриатум оказывает двойственное влияние на нейроны бледного шара - возбуждающее и тормозящее. Электрическое раздражение хвостатого ядра, скорлупы и бледного шара вызывает торможение двигательного компонента условных и безусловных реакций животных. Предполагают, что морфологическим субстратом подобных тормозных влияний являются прямые восходящие связи перечисленных структур с моторной и соматосенсорной корой, а также с таламическими ядрами (вентральными передним и латеральным) и срединным центром.
Головка хвостатого ядра играет важную роль в организации предпусковых процессов, которые включают перестройку позы (см.), предшествующей произвольному двигательному акту. Это подтверждают данные микроэлектродных исследований, выявивших изменения нейрональной активности головки хвостатого ядра в период, предшествующий осуществлению произвольного движения. Так, с помощью микроэлектродных методов Ники (H. Niki с сотр. (1972) показал, что у обезьян в ситуации простого выбора и нажатия на рычаг активация нейронов головки хвостатого ядра перед началом произвольного движения предшествует активации нейронов префронтальной коры. Электромиографическая активность конечности животного регистрируется в среднем через 110 мсек после активации головки хвостатого ядра. По данным Эвартса(E.V. Evarts, 1966), разряд пирамидного нейрона в коре головного мозга обезьяны предшествует мышечной активности ее конечности на 50-100 мсек.
Большинство структур экстрапирамидной системы не имеет прямых выходов к мотонейронам спинного мозга, их влияние на них опосредовано через ретикуло-спинальный тракт, являющийся как бы общим конечным путем экстрапирамидной системы (см. Двигательные центры, пути). Кроме того, экстрапирамидные влияния на спинной мозг и ядра черепно-мозговых нервов осуществляются через трункоспинальные пути, проводящие импульсы от ядер промежуточного мозга (см.), среднего мозга (см.) и продолговатого мозга (см.). Эти пути включают руброспинальный, вестибулоспинальный и оливоспинальный пути. Через эти же пути осуществляется часть мозжечковых влияний на мотонейроны спинного мозга (см. Мозжечок). Общность эффектов этих влияний с экстрапирамидной регуляцией позволяет относить эти структуры к экстрапирамидной системе.
Все супраспинальные структуры, входящие в экстрапирамидную систему, адресуют свои влияния гамма-мотонейронам спинного мозга (см.). Как показали исследования Р. Гранита, гамма-мотонейроны регулируют поток проприоцептивных афферентных импульсов, поступающих в спинной мозг от мышечных веретен (см. Проприоцепторы). Эти афферентные импульсы влияют на возбудимость альфа-мотонейронов, активность которых определяет рабочее состояние скелетных мышц. Например, включение мотонейронов в рефлекс растяжения - миостатический рефлекс - представляет собой часто спинальный процесс, для обеспечения которого достаточно включения одного-двух спинальных сегментов. В этом процессе имеет место прямое управление мотонейронами с помощью сигналов, поступающих от первичных проприорепторов. Нисходящие влияния со стороны структур экстрапирамидной системы могут облегчать или подавлять рефлекс растяжения, что проявляется при децеребрационной ригидности (см.), когда чрезвычайно усиливаются миостатические рефлексы (см. Рефлекс). Фактором, усиливающим их, является увеличение под действием нисходящих влияний активности гамма-мотонейронов, которая, в свою очередь, приводит к возрастанию разрядов рецепторов растяжения и соответствующей интенсификации моносинаптического возбуждения альфа-мотонейронов. Вместе с тем быстропроводящие волокна, берущие начало из медиальной части ретикулярной формации продолговатого мозга и варолиева моста (см. Мост головного мозга), а также из латерального вестибулярного ядра Дейтерса (см. Преддверно-улитковый нерв) тоже моносинаптически возбуждают альфа-мотонейроны спинного мозга и обеспечивают осуществление быстрых движений. Медленнопроводящий нисходящий путь покрышки среднего мозга обеспечивает регуляцию тонических реакций. Таким образом, если нисходящие влияния пирамидной системы (см.), действуя непосредственно на альфа-мотонейроны, повышают их функциональную активность при осуществлении фазных и тонических двигательных реакций организма, то регулирующие влияния экстрапирамидной системы на гамма-мотонейроны обеспечивают необходимую коррекцию выполняемых движений и являются дополнительным механизмом воздействия на позно-тоническую и двигательную активность. Это наиболее выражено при сохранении вертикального положения тела, когда силе тяжести противодействует сокращение мышц-разгибателей, вызываемое облегчающими влияниями стволовых структур экстрапирамидной системы. Тормозные регулирующие влияния, идущие от двигательных центров коры и стриопаллидарных структур, корректируют степень напряжения скелетных мышц.
В настоящее время представления о функциях стриопаллидарных структур существенно расширились. Данные большого количества экспериментальных и клинических исследований свидетельствуют об участии хвостатого ядра, скорлупы, бледного шара не только в регуляции моторной деятельности, но и в анализе афферентных потоков, в регуляции ряда вегетативных функций, в осуществлении сложных форм врожденного поведения, в механизмах кратковременной памяти, а также в регуляции цикла бодрствование - сон. Широкое участие стриопаллидарных образований в организации сложного поведения организмов базируется на обширной мультисенсорной и гетерогенной конвергенции возбуждений к отдельным нервным клеткам. На нейронах происходит взаимодействие афферентных потоков, поступающих практически от всех сенсорных структур, от многих областей коры головного мозга, от таламических, ретикулярных, нигральных, лимбических (см. Лимбическая система) и других структур мозга. Например, при низкой частоте фоновой импульсации и небольшом количестве спонтанно активных нейронов хвостатого ядра почти половина его клеток тем не менее обладает большой конвергентной емкостью и реагирует на звуковой, пищевой и электрокожный раздражители. Взаимодействию нейронов структур экстрапирамидной системы и специфическому включению их в осуществление высших функций способствуют нейротрансмиттеры - дофамин, серотонин, ацетилхолин, ГАМК (см. Гамма-аминомасляная кислота) и др. Подобная химическая гетерогенность синаптических образований в структурах экстрапирамидной системы обеспечивает специализацию включения ее компонентов в механизмы тонкой координации двигательных актов. Избирательное поражение медиаторных процессов в экстрапирамидную систему у человека вызывает появление характерной клинической симптоматики.
Таким образом, широкие афферентные и эфферентные связи структур экстрапирамидной системы между собой, двусторонние связи подкорковых ядер с корой головного мозга, особенно с ее моторными зонами, а также специфические нейромедиаторные связи со структурами промежуточного, среднего и продолговатого мозга обеспечивают широкое взаимодействие в пределах экстрапирамидной системы, что является основой высшей интеграции поведенческих актов и контроля за ними. Функциональная связь экстрапирамидной системы с вегетативными центрами головного мозга обусловливает ее включение в механизмы эмоционально-аффективных реакций организма.
Патология
Поражения экстрапирамидной системы проявляются нарушениями двигательной сферы, при которых не наблюдается клинических признаков поражения пирамидной системы и нарушений чувствительности. Патологические экстрапирамидные синдромы развиваются как при поражении ядер экстрапирамидной системы, так и ее многочисленных связей.
Причиной поражения экстрапирамидной системы могут быть различные заболевания головного мозга - энцефалиты (см.), сосудистые заболевания (см. Атеросклероз , Гипертоническая болезнь), наследственные заболевания центральной нервной системы, черепно-мозговая травма (см.), родовая травма, интоксикации марганцем, окисью углерода, опухоли или гематомы головного мозга глубинной локализации и др. К поражению структур экстрапирамидной системы может привести длительное применение препаратов раувольфин, метилдофа, нейролептических средств (см.), например галоперидола, аминазина и др., а также тяжелая аллергия (см.), асфиксия (см.), полиглобулия (см. Эритроцитозы) и др.
В патогенезе экстрапирамидных синдромов, как было установлено в последние десятилетия, большое значение имеют химические передатчики нервного импульса - медиаторы (см.). При патологии нарушается действие специализированных медиаторов - дофамина, ацетилхолина, гамма-аминомасляной кислоты (ГАМК), серотонина и др., содержащихся в соответствующих нейро-моноаминергических системах мозга (дофаминергических, холинергических, ГАМКергических, серотонинергических и др.). Экстрапирамидные патологические синдромы возникают при дефиците нейроаминов в определенных структурах экстрапирамидной системы, при нарушении нормальной сбалансированности тормозного и облегчающего влияний медиаторных систем. Например, двигательные и эмоциональные проявления паркинсонизма (см.) связаны со снижением активности систем дофаминергических нейронов черного вещества и базальных ядер. Нейромедиаторные нарушения, лежащие в основе хореи Гентингтона (см. Гентингтона хорея), связаны с реципрокным взаимодействием тормозных ГАМК-нейронов и дофаминергических нейронов в полосатом теле. Наследственная дегенерация нейронов первого типа, наблюдающаяся при этом заболевании, способствует высвобождению активности дофаминергической системы и появлению хореических гиперкинезов.
Многообразие экстрапирамидных патологических синдромов обусловлено наличием так наз. анатомо-биохимической диссоциации, выражающейся в том, что морфологические нарушения на одном уровне экстрапирамидной системы приводят с помощью механизма медленного аксонального транспорта нейроаминов к развитию нейрохимических нарушений на другом уровне экстрапирамидной системы, где морфологические патологические изменения отсутствовали.
Для выявления патологии экстрапирамидной системы определяют содержание катехоламинов (см.) и других нейромедиаторов в крови и цереброспинальной жидкости, производят ангиографию (см.), гамма-топографию (см. Энцефалография, радиоизотопная), компьютерную томографию (см. Томография компьютерная), электроэнцефалографию (см.), пневмоэнцефалографию (см.), реоэнцефалографию (см.), исследуют состояние нервно-мышечной системы с помощью методов электромиографии (см.), миотонометрии, стабилографии, треморографии, кимографии (см. Кимография) гиперкинезов в состоянии покоя и при стимуляции и др.
При поражениях экстрапирамидной системы нарушаются двигательные функции, тонус мышц, поза тела, походка, эмоциональные проявления, вегетативнососудистые реакции. Поскольку у человека существует тесная связь между моторикой и мышечным тонусом, при патологии экстрапирамидной системы постоянно встречаются сочетанные нарушения той и другой ее функций. Поражение различных отделов экстрапирамидной системы сопровождается развитием характерных клинических синдромов, которые условно определяют как гипертонически-гипокинетический, связанный преимущественно с патологией паллидума, и гипотонически-гиперкинетический, обусловленный преимущественно патологией стриатума.
Рис. 1. Поза больного паркинсонизмом с акинетико-ригидным синдромом: руки согнуты в лучезапястных и локтевых суставах, ноги полусогнуты в коленных суставах, голова наклонена к груди.
Симптомокомплекс поражения паллидума и его связей характеризуется гипертонически-гипокинетическим статусом, основными проявлениями которого являются повышение мышечного тонуса (см.) и уменьшение двигательной активности (гипокинезия) или практическая обездвиженность при отсутствии параличей (акинезия). Экстрапирамидную мышечную гипертонию (паллидарную мышечную ригидность) называют также восковой, или пластической; при пассивных движениях конечностей больного врач, осуществляющий их, испытывает сопротивление движению, которое остается одинаковым от начала и до конца движения. В случае пассивного разгибания согнутых конечностей больного иногда ощущается прерывистость, своеобразная ступенчатость при пассивном растяжении мышц-сгибателей, названная симптомом зубчатого колеса. Экстрапирамидная гипокинезия обнаруживается на фоне сохранности силы мышц и возможности полного объема движений, которая, однако, не реализуется больным, так как двигательная инициатива его резко снижена. У больных с паллидарным синдромом наблюдается общая скованность с характерной позой: руки согнуты в лучезапястных и локтевых суставах и прижаты к туловищу, ноги полусогнуты в коленных суставах, голова наклонена к груди (так называемый флексорный статус). Больной, приняв ту или иную позу, как бы застывает в этой позе, долго ее сохраняя (рис. 1). Походка замедлена, речь больных глуха, монотонна, без нормальных модуляций. Лицо лишено мимики, маскообразно (см. Мимика). Все произвольные движения совершаются медленно, с трудом (брадикинезия), физиологические синкинезии (см.) отсутствуют, постуральные рефлексы (см.) повышены. Паллидарная мышечная ригидность в сочетании с усилением постуральных рефлексов и поз, выпадением экстрапирамидных кинезов (см. Амиостатический симптомокомплекс) составляет основу синдрома паркинсонизма (см.) и атеросклеротической мышечной ригидности (акинетико-ригидный синдром Ферстера).
Одним из проявлений экстрапирамидных нарушений является дрожание (см.), в патогенезе которого основную роль играют нарушения в системе красное ядро - ретикулярная формация - зубчатое ядро мозжечка. Дрожание (тремор) имеет разную амплитуду движений, частоту, ритмичность и локализацию (тремор пальцев, конечностей, головы и др). Характерное статическое дрожание пальцев рук (тремор покоя) в виде ритмичного движения, напоминающего скатывание пилюль или счет монет, характерно для болезни Паркинсона (см. Дрожательный паралич) и чаще сочетается с экстрапирамидной ригидностью мышц и гипомимией. Статодинамическое, моторное или кинетическое дрожание (появляющееся при движении) является единственным симптомом так наз. эссенциального, идиопатического тремора - самостоятельного хронического, медленно прогрессирующего заболевания экстрапирамидной системы. Кинетический тремор с дрожанием большой амплитуды, возникающим в конечностях, туловище, голове при попытке любого целенаправленного движения, наблюдается при гепато-церебральной дистрофии (см.). К динамическому, интенционному дрожанию относится мозжечковый тип дрожания, который характерен для рассеянного склероза (см.), некоторых энцефалитов.
При поражении ядер покрышки среднего мозга (см.), ретикулярной формации (см.), черной субстанции (см. Средний мозг) появляются экстрапирамидные нарушения в виде фиксированных поз (флексорного или экстензорного статуса) с усилением рефлексов положения (см. Постуральные рефлексы).
Симптомокомплекс поражения стриатума и его связей характеризуется гиперкинетико-гипотоническим статусом, основными проявлениями которого являются разнообразные непроизвольные, насильственные движения, или экстрапирамидные гиперкинезы (см.), вычурные позы, гримасы, жестикуляции, нарушения сложных актов речи, письма, походки, развивающиеся на фоне мышечной гипотонии или дистонии.

Рис. 2. Лицо больного хореей Гентингтона: видна гримаса вследствие хореических гиперкинезов мышц лица.
Обширную группу стриарных нарушений составляют различные виды хореи: малая хорея (см. Ревматизм), хорея Гентингтона (см. Гентингтона хорея), атеросклеротическая хорея, хорея беременных и др. (см. Хорея). Судороги (см.) при хорее быстрые, гиперкинезы размашистые, появляются во всех частях тела, в мышцах лица. Наблюдается гримасничанье (рис. 2), расстраивается речь, письмо, походка становится пританцовывающей. Тонус мышц понижен или изменчив (дистония). После острых нарушений мозгового кровообращения в области полосатого (стриарного) тела и внутренней капсулы может возникнуть синдром гемихореи, при к-ром хореический гиперкинез охватывает лишь половину тела. Н. К. Боголепов (1957) описал так называемый пароксизмальный хореический рубральный гиперкинез, проявляющийся рубящими, размашистыми движениями всех конечностей, сопровождающимися ротацией туловища. В отличие от стриарной хореи в этом случае поражение локализуется в области верхних ножек мозжечка. К вариантам хореического гиперкинеза относят и гемибаллизм (см. Гиперкинезы), при котором наблюдаются бросковые, вращательные движения конечностей одной стороны тела в сочетании с гипотонией мышц. Этот гиперкинез возникает при поражении субталамического ядра и его связей с бледным шаром.

Рис. 3. Рука больного атетозом: гиперкинез в дистальном отделе с переразгибанием пальцев и характерным видом кисти.
Одной из форм экстрапирамидных гиперкинезов является атетоз (см.). Патологические движения в дистальных отделах рук, ног, в лице, шее, наблюдающиеся при атетозе, изменчивы, несинхронны, совершаются как бы с преодолением препятствия, производят впечатление червеобразного, непрерывно текущего спазма (рис. 3).

Рис. 4. Поза больного с торсионным спазмом: голова запрокинута вследствие спастического напряжения мышц шеи.
Мышечный тонус изменчив (дистония). Атетозный гиперкинез характерен для детского церебрального паралича (см. Детские параличи), он бывает также следствием энцефалитов, сосудистых и дегенеративных заболеваний головного мозга. Часто наблюдаются смешанные формы гиперкинезов: хореоатетоз, атетоз с так называемой таламической рукой (см. Таламус) и др. К стриарным экстрапирамидным гиперкинезам относится торсионный спазм. Для него характерны распространенные, чаще вращательные, спазмы больших мышечных масс, вызывающих характерные позы тела с перегибающими (рис. 4), перекручивающими движениями туловища, возникающими при произвольных движениях (см. Торсионная дистония). Торсионный гиперкинез в сочетании с гемибаллизмом, хореическими гиперкинезами, дрожанием (см. выше) и др. наблюдается при гепато-церебральной дистрофии (см.), лейкоэнцефалитах (см.) и других поражениях экстрапирамидной системы.
Тонико-клонические гиперкинезы мышц лица экстрапирамидной природы встречаются при лицевом пара-спазме (спазм Мейжа), охватывающем мышцы верхней части лица, или (при распространенном спазме) все мимические мышцы, а также мышцы шеи и конечностей. Параспазму, как и многим экстрапирамидным синдромам, свойственны парадоксальные кинезин, то есть произвольные установки и позы, к которым больной прибегает для уменьшения или прекращения гиперкинеза.
При поражении экстрапирамидной системы нередко встречаются тонические спазмы взора (см. Взора паралич, судорога), блефароспазм (см.). К локализованным спазмам экстрапирамидного генеза относится спастическая кривошея (см.), при которой шея и голова насильственно фиксированы вследствие спазма мышц шеи в различных позициях (боковой, передней, задней). Этот синдром появляется после энцефалита, интоксикаций. Он обусловлен высвобождением шейно-тонических и лабиринтных рефлексов на уровне оральных отделов мозгового ствола и может сочетаться с другими экстрапирамидными гиперкинезами - дрожанием, торсионной дистонией. Кривошея, обусловленная поражением экстрапирамидной системы, отличается от рефлекторной кривошеи, возникающей при наличии добавочного ребра, при шейном радикулите, остеохондрозе (см. Шейно-плечевые синдромы).
При поражении экстрапирамидной системы может развиваться тик лицевых мышц, мышц брюшной стенки, диафрагмы, гортани (см. Тик). Генерализованный тик в сочетании с «внезапными выкриками» у детей известен как синдром Туретта (см. Туретта синдром). Существует тик диафрагмы, вызывающий икоту и гиперкинезы с респираторными пароксизмами, возникающими в результате сокращения мышц диафрагмы, передней брюшной стенки и характеризующимися приступами быстрых судорожных выдохов, сопровождающихся криками и покашливанием. Во время пароксизма такого респираторного гиперкинеза учащается пульс, наблюдаются вазомоторные расстройства.
Диагноз паллидарных синдромов поражения экстрапирамидной системы с характерными клиническими проявлениями не представляет трудностей. При стриарных гиперкинетических синдромах необходимо многостороннее клиническое обследование для подтверждения органической природы поражения и дифференциальной диагностики с часто наблюдающимися привычными навязчивыми движениями, невротическими тиками, рефлекторными спазмами мышц, не обусловленными органическим поражением экстрапирамидной системы.
Лечение экстрапирамидных поражений направлено прежде всего на основное заболевание. Кроме того, используют патогенетические, симптоматические и общеукрепляющие лекарственные средства. К ним относятся холинолитики, миорелаксанты, бета-блокаторы, средства, содержащие L-ДОФА, фенотиазины. В ряде случаев показаны иглотерапия, аутотренинг. Хирургическое лечение при патологии экстрапирамидной системы заключается в стереотаксических операциях (см. Паллидотомия , Стереотаксическая нейрохирургия , Таламотомия), осуществляемых с целью деструкции отдельных структур экстрапирамидной системы, приводящей к устранению гиперкинезов и уменьшению ригидности мышц.
Библиогр.: Бернштейн Н. А. Очерки по физиологии движений и физиологии активности, М., 1966; Г ранит Р. Основы регуляции движений, пер. с англ., М., 1973; Кандель Э. И. и Войтына С. В. Деформирующая мышечная (торсионная) дистония, М., 1971; Коновалов Н. В. Гепато-церебральная дистрофия, М., 1960; Многотомное руководство по неврологии, под ред. С. Н. Давиденкова, т. 2, с. 133, М., 1962, т. 7, с. 304, 1960; Нейротрансмиттерные системы, под ред. Н. Дж. Легга, пер. с англ., М., 1982; Петелин Л. С. Экстрапирамидные гиперкинезы, М., 1970, библиогр.; Стриопаллидарная система, под ред. Н. Ф. Суворова, Л., 1973; Суворов Н. Ф. Стриарная система и поведение, Л., 1980; Физиологические механизмы движений, под ред. Д. С. Гамбаряна, Ереван, 1978; Частная физиология нервной системы, под ред. П. Г. Костюка, Л., 1983; Шаповалов А. И. Нейроны и синапсы супраспинальных моторных систем, Л., Aldrige J. W., Anderson R. J. a. Murphy J. Т. The role of the basal ganglia in controlling a movement initiated by a visually presented cue, Brain Res., v. 192, p. 3, 1980. H. К. Боголепов, E. И. Минакова;
С. С. Михайлов (ан.), Ю. А. Фадеев (физ.).
Экстрапирамидная система - второй эфферентный механизм реализации произвольных движений и
действий. Экстрапирамидной системой называются все двигательные пути, которые не проходят через
пирамиды продолговатого мозга (рис. 32; цветная вклейка).
Как уже говорилось выше, пирамидная система не является единственным механизмом, с помощью
которого осуществляются произвольные двигательные акты. Экстрапирамидная система более древняя, чем
пирамидная. Она очень сложна по своему составу. До настоящего
времени идут споры о том, какие именно структуры относить к экстрапирамидной системе и, главное,
каковы функции этих структур.
В экстрапирамидной системе различают корковый и подкорковый отделы. К корковому отделу
экстрапирамидной системы относятся те же поля, которые входят в корковое ядро двигательного
анализатора. Это 6-е и 8-е, а также 1-е и 2-е поля, т. е. сенсомоторная область коры. Исключение
составляет 4-е поле, которое является корковым звеном только пирамидной системы. 6-е и 8-е поля по
строению принадлежат к полям моторного типа, в III и V слоях этих полей расположены пирамидные
моторные клетки, но меньшего размера, чем клетки Беца.
Строение подкоркового отдела экстрапирамидной системы довольно сложное. Он состоит из целого ряда
образований. Прежде всего это стриопаллидарная система - центральная группа образований внутри
экстрапирамидной системы, куда входят хвостатое ядро, скорлупа и бледный шар (или паллидум).
Эта система базальных ядер располагается внутри белого вещества (в глубине премоторной зоны мозга) и
характеризуется сложными двигательными функциями (рис. 33).
Следует отметить, что, по современным данным, стриопаллидарная система участвует в осуществлении не
только моторных, но и ряда других сложных функций (см. гл. 22).
В экстрапирамидную систему входят и другие образования: поясная кора, черная субстанция, Льюисово
тело, передневентралъные, интраламинарные ядра таламуса, субталамические ядра, красное ядро,
мозжечок, различные отделы ретикулярной формации, ретикулярные структуры спинного мозга.
Конечной инстанцией экстрапирамидных влияний являются те же мотонейроны спинного мозга, к которым
адресуются импульсы и пирамидной системы (рис. 34).
Четкая анатомическая граница между пирамидной и экстрапирамидной системами отсутствует (рис. 35).
Они обособлены анатомически только на участке пирамид, в продолговатом мозге. Однако функциональные
различия между этими системами достаточно отчетливы. И особенно ясно они проявляются в клинике
локальных поражений головного мозга.
Нарушения двигательных функций, возникающие при различных локальных поражениях мозга, можно
подразделить на относительно элементарные, связанные с поражением исполнительных, эфферентных
механизмов движений, и более сложные, распространяющиеся на произвольные движения и действия и
связанные преимущественно с поражением афферентных механизмов двигательных актов.
Хомская Е. Д. Х = Нейропсихология: 4-е издание. - СПб.: Питер, 2005. - 496 с: ил. 103
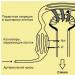
Рис. 33. Стриопаллидум и его эфферентные связи, базальный аспект.
Хвостатое ядро (с. caudatus ), скорлупа (putamen ) и бледный шар (pallidum ), составляющие единую систему (striapallidum ), тесно связаны с глубокими структурами: зрительным бугром, гипоталамической областью, красным ядром, черной субстанцией и корой головного мозга (по О. Криигу )
Относительно элементарные двигательные расстройства возникают при поражении подкорковых звеньев пирамидной и экстрапирамидной систем. При поражении коркового звена пирамидной системы (4-го поля), расположенного в прецентральной области, наблюдаются двигательные расстройства в виде парезов или параличей определенной группы мышц: руки, ноги или туловища на стороне, противоположной поражению. Для поражения 4-го поля характерен вялый паралич (когда мышцы не сопротивляются пассивному движению), протекающий на фоне снижения мышечного тонуса. Но при очагах, расположенных кпереди от 4-го поля (в 6-м и 8-м полях коры), возникает картина спастического паралича, т. е. выпадений соответствующих движений на фоне повышения мышечного тонуса. Явления паре-
Хомская Е. Д. Х = Нейропсихология: 4-е издание. - СПб.: Питер, 2005. - 496 с: ил. 104
Рис. 34. Организация моторных функций на спинальном уровне:
А - латеральный и вентромедиальный пути;
Б - окончание латерального и вентрального путей на мотонейронах передних отделов спинного мозга и на промежуточных
нейронах промежуточной зоны;
В - проекция промежуточных нейронов на мотонейроны и проекция мотонейронов на мускулатуру тела.
Латеральная система проецируется на дистальную мускулатуру, а вентро-медиальная - на проксимальную. 1 -
латеральный кортико-спинальный тракт; 2 - рубро-спинальный тракт; 3 - медиальный тракт; 4 - латеральный ретикуло-
спинальный тракт; 5 - вестибуло-спинальный тракт; 6 - медиальный ретикуло-спинальный тракт; 7 - текто-спинальный
тракт; 8 - вентральный кортико-спинальный тракт; 9 - промежуточная зона; 10 - вентральная зона; 11 -промежуточные
(вставочные) нейроны; 12 - мотонейроны; 13 - пальцы; 14 - руки; 15 - тело (по Л. Лауренсу и Д. Куперсу )
зов вместе с чувствительными расстройствами характерны и для поражения постцентральных отделов коры. Эти нарушения двигательных функций подробно изучаются неврологией. Наряду с этими неврологическими симптомами поражение коркового звена экстрапирамидной системы дает также нарушения сложных произвольных движений, о которых речь будет идти ниже.
Рис. 35. Различные системы афферентаций чувствительных (кинестетических) и двигательных (кинетических) отделов коры (по Д. Пейпецу )
При поражении пирамидных путей в подкорковых областях мозга (например, в зоне внутренней капсулы) возникает полное выпадение движений (паралич ) на противоположной стороне. Полное одностороннее
Хомская Е. Д. Х = Нейропсихология: 4-е издание. - СПб.: Питер, 2005. - 496 с: ил. 105
выпадение движений руки и ноги (гемиплегия ) появляется при грубых очагах. Чаще в клинике локальных поражений мозга наблюдаются явления частичного снижения двигательных функций на одной стороне (гемипарезы ).
При пересечении пирамидного пути в зоне пирамид - единственной зоне, где пирамидный и экстрапирамидный пути анатомически обособлены, - произвольные движения реализуются только с помощью экстрапирамидной системы. В 40-е годы XX века эксперименты по пересечению пирамид проводились английским физиологом С. Тоуэром на обезьянах. Оказалось, что обезьяна способна выполнять целый ряд сложных движений (захватывания, манипуляции с предметами и др.). Однако в подобных случаях движения становятся менее дискретными (т. е. более грубыми). В захватывании предметов начинает участвовать большее количество мышц. Кроме того, в движени-
ях, где требуется участие всех мышц (типа прыжка), что характерно для моторики обезьян, исчезают точность и координированность. Подобные нарушения движений протекают на фоне повышения мышечного тонуса.
Эти данные свидетельствуют о том, что пирамидная система участвует в организации преимущественно точных, дискретных, пространственно-ориентированных движений и в подавлении мышечного тонуса. Поражение корковых и подкорковых звеньев экстрапирамидной системы приводит к появлению различных двигательных расстройств. Эти расстройства можно подразделить на динамические (т. е. нарушения собственно движений) и статические (т. е. нарушения позы). При поражении коркового уровня экстрапирамидной системы (6-е и 8-е поля премоторной коры), который связан с вентролатеральным ядром таламуса, бледным шаром и мозжечком, в контралатеральных конечностях возникают спастические двигательные нарушения. Раздражение 6-го или 8-го полей вызывает повороты головы, глаз и туловища в противоположную сторону (адверзии), а также сложные движения контралатеральных руки или ноги. Поражение подкорковой стриопаллидарной системы, вызванное различными заболеваниями (паркинсонизмом, болезнью Альцгеймера, Пика, опухолями, кровоизлияниями в область базальных ядер и др.), характеризуется общей неподвижностью, адинамией, трудностями передвижения. Одновременно появляются насильственные движения контралатеральных руки, ноги, головы - гиперкинезы. У таких больных наблюдаются и нарушение тонуса (в виде спастичности, ригидности или гипотонии), составляющего основу позы, и нарушение двигательных актов (в виде усиления тремора - гиперкинезов). Больные теряют возможность себя обслуживать и становятся инвалидами.
В настоящее время разработаны хирургические (стереотаксические) и консервативные (фармакологические) методы лечения паркинсонизма. Суть первых состоит в разрушении одной из «больных» экстрапирамидных структур (например, вентролатерального ядра таламуса) и прекращении патологического возбуждения, циркулирующего по замкнутым экстрапирамидным путям. В зависимости от формы паркинсонизма разрушают также либо зону паллидума, либо зону стриатума (в области хвостатого ядра или скорлупы). Зона паллидума (бледный шар) имеет отношение к регуляции медленных плавных движений и к поддержанию тонуса (позы), в то время как зона стриатума (скорлупа и хвостатое ядро) в большей степени
связана с физиологическим тремором, переходящим при патологии в гиперкинезы.
Избирательное поражение зоны паллидума (более древней части, чем стриатум) может привести к атетозу
или хореоатетозу (патологическим волнообразным движениям рук и ног, подергиванию конечностей и т.
д.). Эти заболевания хорошо изучены и описаны в неврологических руководствах.
Поражение стриопаллидарных образований сопровождается еще одним видом двигательных симптомов -
нарушением мимики и пантомимики, т. е. непроизвольных моторных компонентов эмоций. Эти нарушения
могут выступать либо в форме амимии (маскообразное лицо) и общей обездвиженности (отсутствие
непроизвольных движений всего тела при различных эмоциях), либо в форме насильственного смеха, плача
или насильственной ходьбы, бега (пропульсии). Нередко у этих больных страдает и субъективное
"переживание эмоций.
Наконец, у таких больных нарушаются и физиологические синергии - нормальные сочетанные движения
разных двигательных органов (например, размахивание руками при ходьбе), что приводит к
неестественности их двигательных актов.
Последствия поражения других структур экстрапирамидной системы изучены в меньшей степени, за
исключением, конечно, мозжечка. Мозжечок представляет собой важнейший центр координации различных
двигательных актов, «орган равновесия», обеспечивающий целый ряд безусловных моторных актов,
связанных со зрительной, слуховой, кожно-кинестетической, вестибулярной афферентацией. Поражение
мозжечка сопровождается разнообразными двигательными расстройствами (прежде всего расстройствами
координации двигательных актов). Их описание составляет один из хорошо разработанных разделов
современной неврологии.
Поражение пирамидных и экстрапирамидных структур спинного мозга сводится к нарушению функций
мотонейронов, вследствие чего выпадают (или нарушаются) управляемые ими движения. В зависимости от
уровня поражения спинного мозга нарушаются двигательные функции верхних или нижних конечностей (на
одной или на обеих сторонах), причем все местные двигательные рефлексы осуществляются, как правило,
Хомская Е. Д. Х = Нейропсихология: 4-е издание. - СПб.: Питер, 2005. - 496 с: ил. 106
нормально или даже усиливаются вследствие устранения коркового контроля. Все эти нарушения движений также подробно рассматриваются в курсе неврологии.
Клинические наблюдения за больными, у которых имеется поражение того или иного уровня пирамидной или экстрапирамидной системы,
позволили уточнить функции этих систем. Пирамидная система ответственна за регуляцию дискретных, точных движений, полностью подчиненных произвольному контролю и хорошо афферентированных «внешней» афферентацией (зрительной, слуховой). Она управляет комплексными пространственно-организованными движениями, в которых участвует все тело. Пирамидная система регулирует преимущественно фазический тип движений, т. е. движения, точно дозированные во времени и в пространстве.
Экстрапирамидная система управляет в основном непроизвольными компонентами произвольных движений; к ним кроме регуляции тонуса (того фона двигательной активности, на котором разыгрываются фазические кратковременные двигательные акты) относятся:
♦ поддержание позы;
♦ регуляция физиологического тремора;
♦ физиологические синергии;
♦ координация движений;
♦ общая согласованность двигательных актов;
♦ их интеграция;
♦ пластичность тела;
♦ пантомимика;
♦ мимика и т. д.
Экстрапирамидная система управляет также разнообразными двигательными навыками, автоматизмами. В целом экстрапирамидная система менее кортиколизована, чем пирамидная, и регулируемые ею двигательные акты в меньшей степени произвольны, чем движения, регулируемые пирамидной системой. Следует, однако, помнить, что пирамидная и экстрапирамидная системы представляют собой единый эфферентный механизм, разные уровни которого отражают разные этапы эволюции. Пирамидная система, как эволюционно более молодая, является в известной степени «надстройкой» над более древними экстрапирамидными структурами, и ее возникновение у человека обусловлено прежде всего развитием произвольных движений и действий.
(21 Голосов)Нормальное функционирование экстрапирамидной системы и мозжечка у человека – основа, на которой базируется формирование произвольного двигательного акта. Поражение экстрапирамидной системы или мозжечка вызывают разнообразные двигательные нарушения, знание которых является непременным условием в подготовке врача-невролога.
Анатомия и физиология мозжечка.
Мозжечок лежит в задней черепной ямке. Вес мозжечка 120–150 г. Средняя часть мозжечка называется червем. По сторонам от него лежат полушария мозжечка – правое и левое. Параллельными дугообразными бороздами мозжечок разделяется на извилины. Фило- и онтогенетически мозжечок подразделяется на древний (клочок, узелок), старый (червь) и новый (полушария). Мозжечок имеет три пары ножек. Верхняя пара ножек соединяет мозжечок со средним мозгом, средняя – с варолиевым мостом и нижняя – с продолговатым мозгом. Ножки состоят из нервных волокон, которые приносят импульсы к мозжечку или отводят от него. В глубине мозжечка серое вещество образует ядра: зубчатое, пробковидное, шаровидное, а также ядро шатра.
Мозжечок выполняет функцию автоматической координации движений, участвует в регуляции мышечного тонуса и равновесия тела. В осуществлении произвольного движения главная роль мозжечка состоит в согласовании быстрых (фазических) и медленных (тонических) компонентов двигательного акта. Это становится возможным, благодаря двухсторонним связям мозжечка с мышцами и корой головного мозга. Мозжечок получает афферентные импульсы от всех рецепторов, раздражающихся во время движения (от проприоцепторов, вестибулярных, зрительных, слуховых и др.). Получая информацию о состоянии двигательного аппарата, мозжечок оказывает влияние на красные ядра и ретикулярную формацию, которая посылает импульсы к гамма-мотонейронам спинного мозга, регулирующим тонус мышц. Кроме того, часть афферентных импульсов через мозжечок поступает в двигательную зону коры головного мозга.
Однако, основная функция мозжечка, по-видимому, осуществляется на подкорковом уровне (мозговой ствол, спинной мозг). Эфферентные импульсы от ядер мозжечка регулируют проприоцептивные рефлексы на растяжение. Многие симптомы мозжечковой дисфункции связаны с нарушением реципроктной иннервации антагонистов.
Основные афферентные и эфферентные связи мозжечка: путь Флексига (задний спинно-церебеллярный), неперекрещенный; путь Говерса (передний спинно-церебеллярный), дважды совершающий перекрест; лобно-мосто-мозжечковый путь; затылочно-височно-мозжечковый путь.
Существующие перекресты мозжечковых афферентных и эфферентных систем приводят к гомолатеральной связи одного полушария мозжечка и конечностей. Поэтому при поражении полушария мозжечка или боковых столбов спинного мозга мозжечковые расстройства наблюдаются на своей половине тела.
Полушария головного мозга соединены с противоположными гемисферами мозжечка. В связи с этим, при страдании головного мозга или красных ядер мозжечковые расстройства наблюдаются на противоположной половине тела.
В мозжечке существует определенная соматотопика. Считается, что червь мозжечка принимает участие в регуляции мускулатуры туловища, а кора полушарий – дистальных отделов конечностей. Вследствие этого различают статическую и динамическую атаксию.
Симптомы поражения.
А) Статическая атаксия.
Расстраивается в основном стояние и ходьба. Больной стоит с широко расставленными ногами, покачивается. Походка напоминает походку пьяного. Особенно затруднены повороты. В позе Ромберга больной покачивается или вообще не может стоять со сдвинутыми стопами. Это наблюдается как при открытых, так и закрытых глазах. При попытке больного стоя отклониться назад отсутствует наблюдающееся у здоровых людей сгибание в коленных суставах и в поясничном отделе позвоночника. Появляется асинергия (проба Бабинского, синдром отсутствия "обратного толчка" Стюарта-Холмса).
Б) Динамическая атаксия.
Нарушается выполнение различных произвольных движений конечностями. Этот вид атаксии зависит, в основном, от поражения полушарий мозжечка. При выполнении пальценосовой пробы наблюдается промахивание и интенционный тремор. При пяточноколенной пробе больной не попадает пяткой в колено, пятка соскальзывает в сторону при проведении по голени. Промахивание, соскальзывание пятки с голени возникает у больного как с открытыми, так и с закрытыми глазами. Наблюдается адиадохокинез и гиперметрия.
Кроме нарушения движения в конечностях, при поражении мозжечковых систем, расстраиваются и другие простые и сложные двигательные акты: речь (брадилалия, скандированная речь), почерк (мегалография), нистагм. У больных с поражением мозжечка наблюдается также гипотония мышц.
Координация движений нарушается при страдании лобной и височной доли и их проводников. В таких случаях расстраивается ходьба и стояние, туловище отклоняется назад и в сторону, противоположную очагу (астазия-абазия). Выявляется промахивание в руке и ноге – гемиатаксия.
Методика исследования.
Исследование ходьбы, устойчивость в позе Ромберга, проба Бабинского, проба Стюарта-Холмса, координационных проб, диадохокинеза, нарушения речи, наличия нистагма, изменений мышечного тонуса.
Экстрапирамидная система, симптомы поражения.
В обеспечении произвольной моторики человека значительную роль играют многочисленные рефлекторные механизмы, действующие автоматически. Этот большой комплекс нервных структур получил название экстрапирамидной системы. К экстрапирамидной системе относят: бледный шар и полосатое тело, состоящее из скорлупы и хвостатого ядра. Хвостатое ядро со скорлупой составляют вместе неостриатум, в то время как бледный шар является палеостриатумом. Деление на два разных ядра основано как на разновременном появлении этих образований в филогенезе и включении их в действие в онтогенезе, так и на различии в их гистологическом строении. Кроме этого, к экстрапирамидной системе относят субталамические ядра Льюиса, черную субстанцию, красные ядра, зрительные бугры, сетевидное образование, вестибулярные ядра Дейтерса, зубчатое ядро мозжечка, нижние оливы, ядра Даркшевича. В настоящее время к экстрапирамидной системе относят обширные участки коры головного мозга (особенно лобных долей), которые тесно связаны с указанными выше образованиями. Перечисленные составляющие экстрапирамидной системы имеют многочисленные связи. Последние образуют замкнутые нейронные круги, объединяющие многочисленные экстрапирамидные образования ствола и больших полушарий мозга в единые функциональные системы. От коры латеральной, медиальной и нижней поверхностей лобной доли направляются волокна к гомолатеральным ганглиям и ядрам мозгового ствола.
Хвостатое ядро, скорлупа, бледный шар связаны с нижерасположенными клетками мозгового ствола, в частности, с сетевидным образованием. От ядер ствола мозга берут начало пучки волокон, представляющие собой совокупность аксонов соответствующих нервных клеток. Эти пучки проходят в канатиках спинного мозга и заканчиваются синапсами с клетками передних рогов спинного мозга на разных уровнях. К их числу относятся: ретикулоспинальный тракт, оливоспинальный, руброспинальный, тектоспинальный пучки, а также медиальный продольный пучок. Надо полагать, что нисходящие импульсы из черного вещества направляются к мотонейронам спинного мозга, вероятно, через ретикулоспинальный тракт.
Таким образом экстрапирамидная система представляет собой длинную колонку клеток с большим количеством нервных волокон на протяжении всего головного и спинного мозга. Колонка эта местами резко увеличивается в объеме (подкорковые узлы, на некоторых уровнях образуется густое переплетение волокон с телами клеток (бледный шар, сетчатое вещество)).
Открытию функционального значения экстрапирамидной системы способствовали клинико-анатомические наблюдения. Была описана клиническая картина гиперкинеза, гипокинеза, расстройства мышечного тонуса. Ранее была выдвинута концепция, что, гипокинезия зависит от поражения бледного шара, а гиперкинезы связаны с поражением хвостатого ядра и скорлупы. Однако, в последнее время, такой механизм возникновения гипо- и гиперкинеза был отвергнут. Выяснено, что экстрапирамидные расстройства могут возникать как при поражении коры головного мозга, так и при поражении стволового отдела.
Принцип нейронного кольца, замыкающегося при помощи канала обратной связи, признают в настоящее время основным в организации деятельности центральной нервной системы.
Экстрапирамидная система участвует в формировании мышечного тонуса и позы, как бы предуготавливает скелетную мускулатуру в каждое мгновение воспринимать возбудительные и тормозные импульсы. Нарушение в одном из звеньев, регулирующих деятельность экстрапирамидной системы, может привести к ригидности, развитию гипо- или гиперкинеза.
Топический диагноз приходится устанавливать на основании анализа комплекса расстройств различных функций экстрапирамидной системы.
Синдромы поражения экстрапирамидной системы.
1. Паркинсонизм (гипертонический гипокинетический синдром). Для этого синдрома характерны:
- малая двигательная активность больного – олигокинезия (лицо имеет маскообразный вид, взор неподвижен, бедная жестикуляция). Туловище наклонено вперед, руки слегка согнуты в локтевых суставах, прижаты к туловищу. Имеется наклонность застывать в одной, даже неудобной, позе;
- активные движения совершаются очень медленно – брадикинезия (больной ходит мелкими шагами, отсутствуют содружественные движения рук при ходьбе);
- наблюдаются пропульсии;
- мышечная ригидность (симптом "зубчатого колеса");
- наличие гиперкинеза в виде дрожания (ритмичный тремор в пальцах рук напоминающий счет монет или катание пилюль).
В более выраженной форме указанные выше симптомы возникают при очагах в верхних отделах мозгового ствола (вовлечение черной субстанции).
Мышечная ригидность обуславливается недостаточным уровнем дофамина в хвостатом ядре, куда он поступает из черной субстанции. В результате усиливаются облегчающие влияния, идущие из премоторной коры и бледного шара к мотонейронам спинного мозга, что сопровождается повышением тонического рефлекса.
Из других симптомов паркинсонизма необходимо отметить вегетативные расстройства и нарушение психики.
2.Экстрапирамидные гиперкинезы.
А) Хорея (гипотонический гиперкинетический синдром) характеризуется беспорядочными непроизвольными движениями с выраженным локомоторным эффектом, возникает в различных частях тела как в покое, так и во время произвольных двигательных актов. Движения напоминают целесообразные, хотя и утрированные действия. Их сравнивают с пляской, паясничанием. При этом гиперкинезе часто отмечается снижение мышечного тонуса.
Б) Атетоз (неустойчивый) – для этого гиперкинеза характерны медленные тонические сокращения мышц, что внешне похоже на червеобразные движения медленного ритма. Они возникают в покое и усиливаются под влиянием эмоций. Эти периодически наступающие мышечные спазмы, чаще всего локализуются в дистальных отделах рук. Атетоз может быть двусторонним. От хореи, атетоз отличается замедленностью движений и обычно меньшей распространенностью. Иногда в различии этих гиперкинезов возникает затруднение, тогда говорят о хореоатетозе. Атетоз может наблюдаться при поражении различных участков экстрапирамидной системы.
В) Торсионная дистония. У больных, особенно при активных движениях, происходит неправильное распределение тонуса мускулатуры туловища и конечностей. Внешне это выражается тем, что при ходьбе в туловище и конечностях появляются штопорообразные насильственные движения. Торсионно-дистонический гиперкинез может ограничиваться какой-либо частью мышечной системы, например при спастической кривошее. Торсионная дистония возникает при поражении различных участков экстрапирамидной системы (базальные ганглии, клетки мозгового ствола).
Г) Гемибаллизм. Этот редкий тип гиперкинеза локализуется на одной стороне тела, больше страдает рука. В одиночных случаях захватываются обе стороны, тогда говорят о парабаллизме. Проявляется гиперкинез быстрыми размашистыми движениями большого объема, напоминающими бросание или толкание мяча. Эта клиническая картина описывается при поражении Люисова ядра.
Д) Миоклонии – быстрого темпа, обычно беспорядочные сокращения различных мышц или их участков. Небольшая амплитуда, одновременное сокращение антагонистических групп мышц не приводит к выраженному локомоторному эффекту.
Е) Тик – быстрые непроизвольные сокращения мышц. В отличие от функциональных (невротических), тики экстрапирамидного генеза отличаются постоянством и стереотипностью.
Ж) Другие гиперкинезы: лицевой спазм, тоническая судорога взора, миоклонус-эпилепсия.
Красное ядро является основным двигательным координационным центром экстрапирамидной системы. Оно имеет многочисленные связи с корой полушарий большого мозга, со стриопаллидарной системой, с таламусом, с подталамической областью и с мозжечком. Нервные импульсы, поступающие к нейронам красного ядра от коры полушарий, ядер стриопаллидарной системы и ядер промежуточного мозга, после соответствующей обработки следуют по красноядерно-спинномозговому пути, обеспечивающему выполнение сложных привычных движений (ходьба, бег), делая эти движения пластичными, способствуя сохранению определенной позы на протяжении длительного времени, а также обусловливая поддержание тонуса скелетной мускулатуры.
От нейронов полушарий большого мозга, преимущественно из лобной доли, аксоны формируют корково-стриарный тракт, который проходит через переднюю ножку внутренней капсулы. Лишь небольшая часть волокон этого тракта заканчивается непосредственно на мелких мультиполярных клетках красного ядра среднего мозга. Бо́льшая часть волокон направляется к ядрам стриарной системы (базальным ядрам головного мозга), в частности к хвостатому ядру и скорлупе. От нейронов стриарной системы к красному ядру направляется стриарно-красноядерный путь.
Из структур промежуточного мозга с красным ядром связаны нейроны медиальных ядер таламуса (подкорковый чувствительный центр экстрапирамидной системы), нейроны бледного шара (паллидарная система) и нейроны задних ядер гипоталамуса. Аксоны клеток ядер промежуточного мозга собираются в таламо-красноядерный пучок, который заканчивается на клетках красного ядра и черного вещества. Нейроны черного вещества также имеют связи с красным ядром.
Нервные импульсы, поступающие к нейронам красного ядра из мозжечка, осуществляют так называемую поправочную деятельность. Они обеспечивают выполнение тонких целенаправленных движений и предотвращают инерционные проявления при движениях.
Мозжечок связан с красными ядрами посредством двухнейронного пути – мозжечково-красноядерного тракта. Первыми нейронами этого пути являются клетки коры полушарий мозжечка, аксоны которых заканчиваются в зубчатом ядре. Вторыми нейронами являются клетки зубчатого ядра, аксоны которых покидают мозжечок через верхние ножки. Мозжечково-красноядерный тракт входит в средний мозг, па уровне нижних холмиков перекрещивается с одноименным трактом противоположной стороны (перекрест Верне- кинга) и заканчивается на клетках красного ядра (рис. 4.10).
Рис. 4.10.
1 – зубчато-красноядерный путь; 2 – мозжечок; 3 – кора мозжечка; 4 – зубчатое ядро; 5 – шейный сегмент; 6 – поясничный сегмент; 7 – двигательные ядра передних рогов спинного мозга; 8 – красноядерно-спинномозговой путь; 9 – мост; 10 – красное ядро; 11 – средний мозг
От нейронов каждого красного ядра начинается нисходящий красноядерно-спинномозговой путь (пучок Монакова) и красноядерно-ядерный путь, которые сразу же в покрышке среднего мозга переходят на противоположную сторону и образуют передний перекрест покрышки (перекрест Фореля).
Красноядерно-ядерный путь проходит в покрышке ствола головного мозга и заканчивается на мотонейронах двигательных ядер черепных нервов. Аксоны мотонейронов ядер черепных нервов направляются к скелетным мышцам глазного яблока, головы, глотки, гортани и верхней части пищевода, обеспечивая их эфферентную иннервацию.
Красноядерно-спинномозговой путь проходит в боковом канатике спинного мозга. В последнем он располагается кпереди от латерального корково-спинномозгового пути. Постепенно пучок волокон истончается, так как аксоны посегментно заканчиваются на мотонейронах двигательных ядер передних рогов спинного мозга своей стороны. Аксоны мотонейронов покидают спинной мозг в составе передних корешков спинномозговых нервов, а затем в составе самих нервов и их ветвей направляются к скелетным мышцам.
осуществляет безусловнорефлекторные двигательные реакции в ответ па внезапные сильные зрительные, слуховые, тактильные и обонятельные раздражения. Первые нейроны крыше-спинномозгового пути располагаются в верхних холмиках среднего мозга – подкорковом интеграционном центре среднего мозга (рис. 4.11). В данный интеграционный центр информация поступает из подкорковых центров зрения (ядро верхнего холмика), подкоркового центра слуха (ядро нижнего холмика), подкоркового центра обоняния (ядро сосцевидного тела) и коллатералей от проводящих путей общей чувствительности (спинномозговая, медиальная и тройничная петли).
Аксоны первых нейронов направляются вентрально и кверху, обходят центральное серое вещество среднего мозга и переходят на противоположную сторону, образуя задний перекрест покрышки (перекрест Мейнерта). Далее тракт проходит в дорсальной части моста рядом с медиальным продольным пучком. По ходу тракта в стволе головного мозга отходят волокна, которые заканчиваются на мотонейронах двигательных ядер черепных нервов. Эти волокна объединяются под названием крыше-ядерного пучка. Они обеспечивают защитные реакции с участием мышц головы и шеи.
В области продолговатого мозга крыше-спинномозговой путь приближается к дорсальной поверхности пирамид и направляется в передний канатик спинного мозга. В спинном мозге он занимает самую медиальную часть переднего канатика, ограничивая переднюю срединную щель.
Крыше-спинномозговой путь прослеживается на протяжении всего спинного мозга. Постепенно истончаясь, он посегментно отдает ответвления к мотонейронам двигательных ядер передних рогов спинного мозга своей стороны. Аксоны мотонейронов проводят нервные импульсы к мускулатуре туловища и конечностей.

Рис. 4.11.
1 – верхний холмик среднего мозга; 2 – задний перекрест покрышки; 3 – крыше-спинномозговой путь; 4 – двигательные ядра передних рогов спинного мозга; 5 – поясничный сегмент; 6 – шейный сегмент; 7 – продолговатый мозг; 8 – средний мозг
При поражении крыше-спинномозгового тракта исчезают стартовые рефлексы, рефлексы на внезапные звуковые, слуховые, обонятельные и тактильные раздражения.
3. Ретикулярно-спинномозговой путь обеспечивает выполнение сложных рефлекторных актов (дыхательные, хватательные движения и т.д.), требующих одновременного участия многих групп скелетных мышц. Следовательно, он осуществляет координационную роль при этих движениях. Ретикулярно-спинномозговой путь проводит нервные импульсы, оказывающие активирующее или, наоборот, тормозное воздействие на мотонейроны двигательных ядер передних рогов спинного мозга. Кроме того, этот путь передает импульсы, обеспечивающие тонус скелетной мускулатуры.
Первые нейроны ретикулярно-спинномозгового пути располагаются в ретикулярной формации ствола головного мозга. Аксоны этих нейронов идут в нисходящем направлении. В спинном мозге они образуют пучок, который располагается в переднем канатике. Пучок хорошо выражен только в шейном и верхнегрудном отделах спинного мозга. Посегментно он истончается, отдавая волокна к гамма-мотонейронам двигательных ядер передних рогов спинного мозга. Аксоны этих нейронов направляются к скелетным мышцам.
- 4. Преддверно-спинномозговой путь обеспечивает безусловнорефлекторные двигательные акты при изменении положения тела в пространстве. Преддверно-спинномозговой путь образован аксонами клеток латерального и нижнего вестибулярных ядер (ядер Дейтерса и Роллера). В продолговатом мозге он располагается в дорсальном отделе. В спинном мозге проходит на границе бокового и переднего канатиков, поэтому пронизан горизонтально ориентированными волокнами передних корешков спинномозговых нервов. Волокна преддверно-спинномозгового пути посегментно заканчиваются на мотонейронах двигательных ядер передних рогов спинного мозга. Аксоны мотонейронов в составе корешков спинномозговых нервов покидают спинной мозг и направляются к скелетной мускулатуре, обеспечивая перераспределение тонуса мышц в ответ на изменение положения тела в пространстве.
- 5. Оливо-спинномозговой путь обеспечивает безусловнорефлекторное поддержание тонуса мышц шеи и двигательные акты, направленные па сохранение равновесия.
Оливо-спинномозговой путь начинается от нейронов нижнего оливного ядра продолговатого мозга. Являясь филогенетически новым образованием, нижнее оливное ядро имеет непосредственные связи с корой полушарий лобной доли (корково-оливный путь), с красным ядром (красноядерно-оливный путь) и с корой полушарий мозжечка (оливо-мозжечковый путь). Аксоны клеток нижнего оливного ядра собираются в пучок – оливо-спинномозговой путь, который проходит в переднемедиальном отделе бокового канатика. Он прослеживается только на уровне шести верхних шейных сегментов спинного мозга.
Волокна оливо-спинномозгового тракта посегментно заканчиваются на мотонейронах двигательных ядер передних рогов спинного мозга, аксоны которых в составе передних корешков спинномозговых нервов покидают спинной мозг и направляются к мышцам шеи.
6. Медиальный продольный пучок осуществляет согласованные движения глазных яблок и головы. Эта функция необходима для поддержания равновесия тела. Выполнение данной функции становится возможным только в результате морфофункциональной связи между нервными центрами, обеспечивающими иннервацию мышц глазного яблока (двигательные ядра III, IV и VI пар черепных нервов), центрами, отвечающими за иннервацию мышц шеи (двигательное ядро XI пары и двигательные ядра передних рогов шейных сегментов спинного мозга), центром равновесия (ядро Дей- терса). Координируют работу названных центров нейроны крупных ядер ретикулярной формации – интерстициального ядра (ядра Кахаля) и ядра задней спайки (ядра Дарк- шевича).
Интерстициальное ядро и ядро задней спайки располагаются в ростральном отделе среднего мозга в его центральном сером веществе. Аксоны нейронов этих ядер формируют медиальный продольный пучок, который проходит под центральным серым веществом вблизи срединной линии. Не меняя своего положения, он продолжается в дорсальной части моста и в вентральном направлении отклоняется в продолговатом мозге. В спинном мозге он располагается в переднем канатике, в углу между медиальной поверхностью переднего рога и передней белой спайкой. Прослеживается медиальный продольный пучок только на уровне верхних шести шейных сегментов.
От медиального продольного пучка направляются волокна к двигательному ядру глазодвигательного нерва, иннервирующему большинство мышц глазного яблока. Далее в пределах среднего мозга из состава медиального продольного пучка направляются волокна к нейронам двигательного ядра блокового нерва стороны. Это ядро отвечает за иннервацию верхней косой мышцы глазного яблока.
В мосту в состав медиального продольного пучка вступают аксоны клеток ядра Дейтерса (VIII пара), которые идут в восходящем направлении к нейронам интерстициального ядра. От медиального продольного пучка отходят волокна к нейронам двигательного ядра отводящего нерва (VI пара), отвечающего за иннервацию латеральной прямой мышцы глазного яблока. И, наконец, в пределах продолговатого и спинного мозга от медиального продольного пучка волокна направляются к нейронам двигательного ядра добавочного нерва (XI пара) и двигательным ядрам передних рогов шести верхних шейных сегментов, отвечающих за функцию мышц шеи.
Кроме общей координации работы мышц глазного яблока и головы, медиальный продольный пучок выполняет важную интегративную роль в деятельности мышц глаза. Осуществляя связь с клетками ядра глазодвигательного и отводящего нервов, он обеспечивает согласованную функцию наружной и внутренней прямых мышц глаза, проявляющуюся в сочетанном повороте глаз в сторону. При этом происходит одновременное сокращение латеральной прямой мышцы одного глаза и медиальной прямой мышцы другого глаза.
При поражении интерстициального ядра или медиального продольного пучка происходит нарушение координированной работы мышц глазного яблока. Чаще всего это проявляется в виде нистагма (частые сокращения мышц глазного яблока, направленные в сторону движения, при остановке взгляда). Нистагм может быть горизонтальным, вертикальным и даже ротаторным (вращательным). Нередко указанные нарушения дополняются вестибулярными расстройствами (головокружение) и вегетативными расстройствами (тошнота, рвота и т.д.).
7. Задний продольный пучок осуществляет связи между вегетативными центрами ствола головного и спинного мозга.
Задний продольный пучок (пучок Шютце) берет начало от клеток задних ядер гипоталамуса. Аксоны этих клеток объединяются в пучок лишь на границе промежуточного и среднего мозга. Далее он проходит в непосредственной близости от водопровода среднего мозга. Уже в среднем мозге часть волокон заднего продольного пучка направляется к добавочным ядрам глазодвигательного нерва. В области моста от заднего продольного пучка отходят волокна к слезному и верхнему слюноотделительному ядрам лицевого нерва. В продолговатом мозге ответвляются волокна к нижнему слюноотделительному ядру языкоглоточного нерва и дорсальному ядру блуждающего нерва.
В спинном мозге задний продольный пучок располагается в виде узкой ленты в боковом канатике, рядом с латеральным корково-спинномозговым трактом. Волокна пучка Шютце посегментно заканчиваются на нейронах промежуточно-латеральных промежуточных ядер, являющихся вегетативными симпатическими центрами спинного мозга.
Лишь небольшая часть волокон заднего продольного пучка обособляется на уровне поясничных сегментов и располагается вблизи центрального канала. Этот пучок носит название околоэпендимального. Волокна данного пучка заканчиваются на нейронах крестцовых парасимпатических ядер.
Аксоны клеток парасимпатических и симпатических ядер покидают ствол головного или спинного мозга в составе черепных или спинномозговых нервов и направляются к внутренним органам, сосудам и железам. Таким образом, задний продольный пучок играет очень важную интегративную роль в регуляции жизненно важных функций организма.
б) Экстрапирамидные пути. Подразделение двигательной системы на пирамидную и экстрапирамидную является источником путаницы и ошибок, так как вначале пирамидную систему представляли единственной двигательной системой. Вероятно, поэтому все двигательные пути, описанные позднее, были объединены в экстрапирамидную систему. Провести четкую грань в анатомическом строении и функциональном значении между пирамидной и экстрапирамидной системами практически невозможно, так как они функционируют в полном взаимодействии. С помощью экстрапирамидной системы тело поддерживается в нужном положении и выполняются содружественные движения с регуляцией мышечного тонуса при выполнении безусловнорефлекторных двигательных актов.
К экстрапирамидной системе относятся многие рассеянные двигательные клетки среди чувствительной зоны, базальные ядра (бледный шар, скорлупа, хвостатое ядро, ограда), передненижнее ядро таламуса, субталамические ядра, мозжечок, красное ядро, черная субстанция и ядра ретикулярной формации.
Экстрапирамидная проводящая система начинается от разбросанных по всей коре головного мозга двигательных клеток, аксоны которых поступают через заднюю ножку внутренней капсулы к хвостатому ядру и скорлупе, а затем после переключения идут к бледному шару. От наружного членика бледного шара аксоны направляются в черное вещество, красное ядро, медиальное вестибулярное ядро и ретикулярную формацию.
От ядер ретикулярной формации берет начало ретикулярно-спинномозговой путь (tr. reticulospinalis), вступающий в контакт со вставочными нейронами и малыми альфа-нейронами передних столбов спинного мозга. От красных ядер формируется tr. rubrospinalis, входящий в боковой канатик спинного мозга и контактирующий со вставочными нейронами заднего и переднего столбов спинного мозга, которые переключаются на малые альфа- и гамма-нейроны.
В экстрапирамидной системе имеется замкнутая петля проводящих путей; она начинается в бледном шаре и проходит к таламусу, где есть центр «активации», затем к скорлупе и вновь возвращается к бледному шару. Бледный шар связан с моторными клетками коры, которые и активируются под действием возвратных импульсов, посылая прямые сигналы к спинному мозгу через пирамидные пути. Допускается, что полосатые тела (хвостатое ядро, скорлупа, бледный шар) являются первичными инициаторами двигательной активности после получения сигналов из сенсорных областей. Кора не доминирует над полосатым телом, но в процессе формирования движений повышает способность экстрапирамидной системы к включению в рефлекторный процесс и увеличивает ее гибкость в процессе перестройки.